En el siguiente enlace hace referencia a los medicamenentos que contribuyen a la una mejor calidad en la salud del ser humano .
http://www.info-farmacia.com/bioquimica/metabolismo-de-farmacos-enzimas-cyp3a-sub-familia-del-citocromo-p450
miércoles, 6 de julio de 2016
El agua y los aminoacidos:
El agua es el elemento más importante para la vida en la Tierra. El 70 por ciento del planeta está formado por agua. A su vez, el ser humano y la mayoría de animales están constituidos por un 70 por ciento de agua, y las propias células de nuestro interior, en un 70 por ciento están compuestas por agua. Este dato ya nos está poniendo en antecedentes sobre la importancia biológica que tiene el agua para la vida y para los seres vivos que habitamos la Tierra.
El agua, una molécula de estructura simple pero de comportamiento extraño, puede ser considerada como el líquido de la vida: los organismos vivos dependen absolutamente de ella para su existencia. No sólo es la molécula más abundante en la biosfera, donde se la encuentra en sus tres estados, sino que además es el componente mayoritario de los seres vivos, en los cuales constituye entre el 65 y el 95% de la masa de las células y de los tejidos. El agua es, entonces, el componente esencial de la estructura y de la función de los tejidos de los seres vivos.
El agua es también considerada el soporte de donde surgió la vida. Sus propiedades físicas y químicas la hacen diferente a la mayoría de los líquidos y son las que le confieren su importancia biológica. Durante la evolución de la vida, los organismos se han adaptado al ambiente acuoso y han desarrollado sistemas que les permiten aprovechar las inusitadas propiedades del agua.
El agua, respecto a otros líquidos, resulta ser un líquido extraño que tiene unas características especiales obtenidas de su propia composición molecular. El agua esta constituida por dos átomos de hidrogeno y uno de oxigeno, mas conocida como H2O.
A continuacion un resumen que se trata sobre la generalidades de un aminoacido:
El agua, una molécula de estructura simple pero de comportamiento extraño, puede ser considerada como el líquido de la vida: los organismos vivos dependen absolutamente de ella para su existencia. No sólo es la molécula más abundante en la biosfera, donde se la encuentra en sus tres estados, sino que además es el componente mayoritario de los seres vivos, en los cuales constituye entre el 65 y el 95% de la masa de las células y de los tejidos. El agua es, entonces, el componente esencial de la estructura y de la función de los tejidos de los seres vivos.
El agua es también considerada el soporte de donde surgió la vida. Sus propiedades físicas y químicas la hacen diferente a la mayoría de los líquidos y son las que le confieren su importancia biológica. Durante la evolución de la vida, los organismos se han adaptado al ambiente acuoso y han desarrollado sistemas que les permiten aprovechar las inusitadas propiedades del agua.
El agua, respecto a otros líquidos, resulta ser un líquido extraño que tiene unas características especiales obtenidas de su propia composición molecular. El agua esta constituida por dos átomos de hidrogeno y uno de oxigeno, mas conocida como H2O.
A continuacion un resumen que se trata sobre la generalidades de un aminoacido:
¿Qué es un aminoácido?
Todos los aminoácidos comparten una estructura química común. Un grupo de amino (representado químicamente como NH2) está unido a un átomo de carbono (el carbono central o alfa) que después se une a otro átomo de carbono. Éste se encuentra en la forma de ácido carboxílico (abreviatura química COOH). El grupo de amino y el grupo de ácido carboxílico tienen una participación crucial en los enlaces que se forman entre los aminoácidos cuando se sintetizan las proteínas.
¿Qué diferencias hay entre los distintos aminoácidos? Los aminoácidos son diferentes en virtud de la presencia de diferentes grupos químicos unidos al átomo de carbono alfa, lo que se conoce comúnmente como cadenas laterales.
Los aminoácidos y el agua o la grasa
Dependiendo de la cadena lateral que esté presente en el carbono alfa, un aminoácido en particular puede tener un comportamiento químico muy diferente. Debido a la naturaleza química de las diferentes cadenas laterales que se utilizan, hay grupos de aminoácidos que no les afecta ser sumergidos en agua, éstos son los aminoácidos hidrofílicos. Los aminoácidos hidrofílicos, pueden sumergirse en el ambiente acuoso del interior de la célula, sin ver su naturaleza afectada. También existen aquellos aminoácidos que no se detectan en el agua, sino que prefieren ocultarse en la grasa o en las suspensiones oleosas. Éstos son los aminoácidos hidrófobos, como los que se encuentran en los confines de las membranas celulares.
Entre algunos ejemplos de aminoácidos hidrófilos (“amantes del agua”) podemos incluir la lisina, la arginina, el ácido aspártico y el ácido glutámico. Entre los ejemplos de aminoácidos hidrófobos (“temor al agua”) podemos nombrar la leucina, la isoleucina, la fenilalanina y la valina.
La síntesis de las proteínas a partir de los aminoácidos
Las proteínas son polímeros lineales de aminoácidos. Las instrucciones que están codificadas en nuestros genes especifican el orden en que los aminoácidos específicos deben unirse entre sí para formar una proteína en particular, tal como la insulina. El primer aminoácido en la cadena dona parte de su grupo de ácido carboxílico para formar parte de un enlace químico con el grupo de amino del aminoácido siguiente en la cadena y así, sucesivamente a medida que el polímero se sintetiza. Cuando se termina una cadena, el primer aminoácido todavía tiene un grupo de amino no utilizado, por lo que se conoce como el amino terminal. Del mismo modo, el último aminoácido de la cadena tiene un grupo ácido carboxílico no utilizado y por lo tanto, el final de la proteína se conoce como carboxilo terminal.
Los aminoácidos son necesarios en nuestra dieta todos los días. Las células humanas pueden sintetizar 10 aminoácidos. Los otros 10 aminoácidos restantes utilizados habitualmente, debemos adquirirlos a través de nuestra dieta. Éstos son los llamados aminoácidos esenciales, entre ellos podemos nombrar la arginina, la histidina la lisina, la metionina, la isoleucina, la leucina, la fenilalanina, la valina, la treonina y el triptófano. Necesitamos todos estos aminoácidos no sólo con el fin de crear las proteínas celulares que nuestro cuerpo requiere para su buen funcionamiento, sino también para la síntesis de otros compuestos e incluso, en casos seleccionados, para utilizarlos como señales del sistema nervioso.
jueves, 30 de junio de 2016
ACIDOS NUCLEICOS
Los ácidos nucleicos son grandes polímeros formados por la repetición de monómeros denominados nucleótidos, unidos mediante enlaces fosfodiéster. Se forman, así, largas cadenas; algunas moléculas de ácidos nucleicos llegan a alcanzar tamaños gigantescos, con millones de nucleótidos encadenados.





A continuacion tres videos recomendados por la profesora en teoria
a) Iniciación. Comienza por el triplete iniciador del ARNm (AUG), que está próximo a la caperuza 5'. Este triplete va precedido de la secuencia AGGAGG (secuencia de Shine-Dalgarno ) que es la zona de unión con el ribosoma.
Se forma el complejo de iniciación con los factores de iniciación (FI) y la energía suministrada por el GTP, la subunidad menor del ribosoma reconoce la caperuza y se une al ARNm en la zona proxima al triplete o codón iniciador. Esta caperuza aporta el ARNt iniciador que a su vez aporta el aminoácido metionina. Este ARNt contiene un triplete complementario al AUG, es decir el UAC, llamado anticodón (la proteína sintetizada contiene en su extremo el aminoácido metionina)
Una vez encajado el ARNt-metionina, se liberan los FI y dejan paso a la subunidad mayor del ribosoma, formandose así el ribosoma completo y funcional. En él hay dos sitios claves:
- Sitio P (sitio peptidil) ocupado por el ARNt-metionina
- Sitio A (sitio aminoacil) que está libre para recibir un segundo ARNt (sólo el que su anticodón coincida con el del codón del ARNm) cargado con un nuevo aminoácido.

 b) Elongación de la cadena peptídica: es un proceso catalizado por el enzima peptidil transferasa, el cual, mediante enlaces peptídicos va uniendo aminoácidos a la cadena peptídica. Cada vez que llega un aminoácido ocurre un proceso cíclico de elongacion.
b) Elongación de la cadena peptídica: es un proceso catalizado por el enzima peptidil transferasa, el cual, mediante enlaces peptídicos va uniendo aminoácidos a la cadena peptídica. Cada vez que llega un aminoácido ocurre un proceso cíclico de elongacion.

 c) Fin de la síntesis de la cadena peptídica: ocurre cuando aparece uno de los codones de terminación ( UAA,UAG,UGA ). En este momento un factor proteico de terminación (RF) se une al codón de terminación e impide que algún ARNt con otro aminoácido (ARNt-aminoacil) se aloje en el sitio A. En este momento se produce la hidrólisis de la cadena peptídica y se separan las dos subunidades del ribosoma.
c) Fin de la síntesis de la cadena peptídica: ocurre cuando aparece uno de los codones de terminación ( UAA,UAG,UGA ). En este momento un factor proteico de terminación (RF) se une al codón de terminación e impide que algún ARNt con otro aminoácido (ARNt-aminoacil) se aloje en el sitio A. En este momento se produce la hidrólisis de la cadena peptídica y se separan las dos subunidades del ribosoma.

Los ácidos nucleicos almacenan la información genética de los organismos vivos y son los responsables de la transmisión hereditaria. Existen dos tipos básicos, el ADN y el ARN.
El descubrimiento de los ácidos nucleicos se debe a Friedrich Miescher, quien en el año 1869 aisló de los núcleos de las células una sustancia ácida a la que llamó nucleína,1 nombre que posteriormente se cambió a ácido nucleico.
Posteriormente, en 1953, James Watson y Francis Crick descubrieron la estructura del ADN, empleando la técnica de difracción de rayos X.
ESTRUCTURAS ADN
- Estructura primaria. Una cadena de desoxirribonucleótidos (monocatenario) es decir, está formado por un solo polinucleótido, sin cadena complementaria. No es funcional, excepto en algunos virus.
- Estructura secundaria. Doble hélice, estructura bicatenaria, dos cadenas de nucleótidos complementarias, antiparalelas, unidas entre sí por medio de las bases nitrogenadas por medio de puentes de hidrógeno.
Está enrollada helicoidalmente en torno a un eje imaginario.
Hay tres tipos:
- Doble hélice A, con giro dextrógiro, pero las vueltas se encuentran en un plano inclinado (ADN no codificante).
- Doble hélice B, con giro dextrógiro, vueltas perpendiculares (ADN funcional).
- Doble hélice Z, con giro levógiro, vueltas perpendiculares (no funcional); se encuentra presente en los parvovirus.
CLASIFICACIÓN
Nucleósidos y nucleótidos
Las unidades que forman los ácidos nucleicos son los nucleótidos. Cada nucleótido es una molécula compuesta por la unión de tres unidades: un monosacárido de cinco carbonos (una pentosa, ribosa en el ARN y desoxirribosa en el ADN), una base nitrogenada purínica (adenina, guanina) o pirimidínica (citosina, timina o uracilo) y un grupo fosfato (ácido fosfórico).
Tanto la base nitrogenada como los grupos fosfato están unidos a la pentosa.
La unidad formada por el enlace de la pentosa y de la base nitrogenada se denomina nucleósido.
El conjunto formado por un nucleósido y uno o varios grupos fosfato unidos al carbono 5′ de la pentosa recibe el nombre de nucleótido.
Se denomina nucleótido-monofosfato (como el AMP) cuando hay un solo grupo fosfato, nucleótido-difosfato (como el ADP) si lleva dos y nucleótido-trifosfato (como el ATP) si lleva tres.
PROPIEDADES Listado de las bases nitrogenadas
Las bases nitrogenadas conocidas son:
- Adenina, presente en ADN y ARN
- Guanina, presente en ADN y ARN
- Citosina, presente en ADN y ARN
- Timina, presente exclusivamente en el ADN
- Uracilo, presente exclusivamente en el ARN
A continuacion tres videos recomendados por la profesora en teoria
Replicación de ADN
Los eventos de la replicación dan como resultado moléculas iguales
de ADN, Este proceso de replicación es semiconservativo ya que se necesita la
cadena parental (cadena molde), para obtener de esta manera un nueva cadena
complementaria.
Como primer punto, el ADN ocurre el rompimiento de los
puentes de hidrógenos que se encuentran entre las dos cadenas existentes, esto
ocurre gracias a la enzima helicaza, luego la proteína SSBS no permite que
estas cadenas separadas se vuelvan a unir, este proceso crea una burbuja de replicación,
que se observa a lo largo de la molécula de ADN, este proceso aumenta la
velocidad de la replicación.
Una vez formada la burbuja de replicación se observara con detalle
a la horquilla de replicación, la ADN polimerasa comienza a formar otra cadena nueva,
luego se separan y se desenrollan hebra conductora es la nueva cadena que se
crea de modo continuo.
La ADN polimerasa construye la nueva cadena EN DIRECCION 5” A
3”, la ADN prolonga una cadena preexistente, ARN polimerasa coloca los primeros
nucleótidos de la cadena.
El segmento resultante ARN cebados proporciona un extremo 3”
libre que se puede enlazar, el ADN coloca los nucleótidos a lo largo del molde.
Luego un tipo diferente de ADN polimerasa reemplaza el ARN cebador
el ARN por ADN, en este punto se produce la polimerización que es el proceso
por el cual se forman las nuevas cadenas, el fosfato se un al OH libre y se forman
puentes de hidrógenos entre los nucleótidos.
La hebra rezagada se sintetiza en dirección opuesta del
avance de la horquilla, se presentan los fragmentos de okazaki.
Transcripción
La transcripción es el proceso por el cual se sintetiza un ARN usando como molde al ADN. Muchos tipos de ARN pueden ser sintetizados asì por la enzima ARN polimerasa, el ARN ribosomal el de transferencia, los pequeños ARN nucleares o citoplasmáticos y por supuesto los ARN mensajeros, que serán luego traducidos a una cadena polipeptídica. El proceso de la transcripción de los mensajeros es diferente en procariotas y eucariotas. Esto es debido a las diferencias propias entre los genes de las bacterias y los de las celulas de animales superiores.
La transcripción es el proceso de obtención de un ARN mensajero (ARNm) a partir del ADN correspondiente a un gen. En cada tipo celular y en cada estado funcional en el que se encuentre la célula se expresan unos genes diferentes. Esto precisa una regulación muy fina que consiga orquestar un funcionamiento complejo de las células orientado a responder adecuadamente a los estímulos externos e internos.
Primero se lleva a cabo la copia de ADN y se obtiene pre-ARNm. Se transcriben tanto los exones (parte codificante de proteína) como los intrones (parte no codificante). La copia de ADN a pre-ARNm se produce nucleótido a nucleótido, incorporando uracilo como base complementaria a la adenina. Como molde se usa una sola cadena de ADN, obteniéndose por tanto pre-ARNm monocatenario. En el núcleo el pre-ARNm sufre un proceso de maduración y una vez maduro sale al citosol donde se traduce a proteína gracias a los ribosomas.
La transcripción la realiza la ARN polimerasa II. La ARN polimerasa II es la principal responsable de copiar la información de los genes pero necesita la participación de muchos otros factores (factores de transcripción, acetilasas de histonas, complejo remodelador de la cromatina) para llevar a cabo este complejo proceso clave en la fisiología celular. El proceso de transcripción se realiza en varias fases:
• Fase de iniciación de la transcripción. Para que se inicie la transcripción es necesario que el gen esté accesible para lo que la cromatina debe disminuir su grado de empaquetamiento. La acetilación de determinadas lisinas de las histonas favorece la descondensación de la cromatina. El complejo remodelador de la cromatina o CRM (Chromatin-Remodeling-Machine) tiene afinidad por las lisinas acetiladas de las histonas y se une a ellas a través del dominio llamado "Bromodominio". El CRM desorganiza los nucleosomas aumentando el grado de exposición y de accesibilidad de los promotores. Finalmente el mediador, la ARN polimerasa II y los factores de transcripción hacen que se inicie la transcripción.
El promotor de un gen es una región del ADN con unas características especiales que determina el punto en el que la ARN polimerasa comienza a transcribir un gen. Una vez que la ARN polimerasa ha reconocido el promotor y se ha unido a él en primer lugar se forma el complejo de preiniciación. Este complejo está formado por algunos factores de transcripción y la ARN polimerasa II. Después se forma el complejo de iniciación cerrado al unirse otros factores de transcripción y el mediador. Posteriormente se forma el complejo de iniciación abierto gracias a la actividad helicasa de uno de los factores. En este instante comienza la síntesis de ARNm por la ARN polimerasa II a partir del sitio llamado +1 que marca el punto de inicio de la transcripción de un gen. Hasta que el fragmento de ARNm sintetizado no tiene 8-10 nucleótidos el proceso es reversible siendo posible que el complejo se desorganice y la transcripción del gen no se lleve a cabo. A este fenómeno se le conoce como iniciación abortiva. Cuando ya hay un fragmento de ARN de tamaño adecuado, se fosforila el dominio CTD (Carboxi-Terminal Domain) de la ARN polimerasa II. Esta fosforilación, llevada a cabo por el mismo factor que tiene actividad helicasa o por el mediador, es muy importante, ya que desestabiliza las interacciones que tiene la ARN polimerasa II con algunos factores de transcripción favoreciendo su avance rápido transcribiendo el gen.
• Fase de elongación. Durante la fase de elongación el CTD debe seguir fosforilado. En esta etapa, la ARN polimerasa II cataliza la formación de los enlaces fosfodiéster entre nucléotidos. Intervienen otros factores, conocidos como factores de elongación. Sus funciones son disminuir las pausas de la ARN polimerasa II, desorganizar los nucleosomas y favorecer los procesos de corrección de errores. También intervienen factores de transcripción involucrados en la iniciación. En la elongación se fosforila otra posición del CTD. El CTD fosforilado es reconocido también por proteínas encargadas del procesamiento y maduración del pre-ARNm, de modo que la maduración del pre-ARNm se produce de forma simultánea a la transcripción. Concretamente, la formación de la caperuza (cap) metilada ocurre en la fase de iniciación de la transcripción y normalmente, el proceso de splicing durante la elongación. La poliadenilación comienza en la fase de terminación de la transcripción y acaba una vez finalizada ésta.
• Fase de terminación. El CTD es defosforilado por una fosfatasa. La ARN polimerasa II continúa transcribiendo hasta llegar a una secuencia específica que determina el sitio de poliadenilación. Esta secuencia es reconocida por una endonucleasa. Esta enzima corta el ARNm unos nucleótidos más allá de la secuencia que reconoce, liberándolo. Posteriormente esta secuencia específica es reconocida por una enzima encargada de añadir la cola de poli-A al transcrito.
El proceso de transcripción debe estar finamente regulado, ya que de él depende la adaptación al medio y el buen funcionamiento del organismo. Cada tipo celular y cada estado funcional necesita un perfil de expresión génica diferente que se adapte a sus necesidades. Los factores reguladores pueden incidir sobre cualquiera de los elementos de este complejo proceso. Así hay reguladores que modifican la accesibilidad de los genes, otros que alteran la fase de iniciación, otros que actúan en la fase de elongación y otros en la de terminación. Igualmente existen reguladores de la transcripción que actúan sobre los procesos de maduración del ARN
La regulación de la accesibilidad de los genes depende de las regiones controladoras de locus o LCRs (Locus Control Region), de los “insulators” (aisladores), de los procesos de modificación de histonas, de los procesos de desempaquetamiento y desorganización de nucleosomas y de la metilación del ADN:
• Las LCRs son regiones del ADN donde la expresión génica está favorecida.
• Los “insulators” son regiones del ADN que delimitan a modo de barrera las zonas transcripcionalmente activas de las que están inactivas participando en la correcta organización de la cromatina.
• Las modificaciones en las histonas son muy importantes en el control de la transcripción. La metilación de lisinas impide la transcripción génica ya que las histonas con este tipo de modificación reclutan proteínas silenciadoras de genes que suelen poseer un “cromodominio” que intervienen en la interacción con las histonas metiladas. En cambio, la acetilación de determinadas lisinas favorece la interacción con el CRM que lleva a cabo la desorganización de los nucleosomas y por tanto hace los genes accesibles para ser transcritos.
• La metilación del ADN es otra modificación que produce silenciamiento de genes. La metilación de genes es muy importante en procesos del desarrollo, en la impronta ganética y en el silenciamiento de genes repetidos.
Otro nivel de regulación de la transcripción es el que se lleva a cabo actuando sobre el proceso de iniciación. Existen proteínas activadoras que se unen a elementos próximos al promotor y otras que se unen a los llamados “potenciadores” que pueden localizarse en posiciones más alejadas del promotor. Las proteínas activadoras se unen a sus sitios de unión en el ADN y favorecen el reclutamiento de la ARN polimerasa II y de los factores de transcripción necesarios para que se forme el complejo de iniciación y comience la transcripción. Las proteínas represoras actúan de forma directa uniéndose al operador y evitando que se una la ARN polimerasa o de forma indirecta bloqueando la acción de las proteínas activadoras.
Otros reguladores de la transcripción actúan sobre las proteínas involucradas en el proceso de elongación bloqueando la transcripción a este nivel.
Primero se lleva a cabo la copia de ADN y se obtiene pre-ARNm. Se transcriben tanto los exones (parte codificante de proteína) como los intrones (parte no codificante). La copia de ADN a pre-ARNm se produce nucleótido a nucleótido, incorporando uracilo como base complementaria a la adenina. Como molde se usa una sola cadena de ADN, obteniéndose por tanto pre-ARNm monocatenario. En el núcleo el pre-ARNm sufre un proceso de maduración y una vez maduro sale al citosol donde se traduce a proteína gracias a los ribosomas.
La transcripción la realiza la ARN polimerasa II. La ARN polimerasa II es la principal responsable de copiar la información de los genes pero necesita la participación de muchos otros factores (factores de transcripción, acetilasas de histonas, complejo remodelador de la cromatina) para llevar a cabo este complejo proceso clave en la fisiología celular. El proceso de transcripción se realiza en varias fases:
• Fase de iniciación de la transcripción. Para que se inicie la transcripción es necesario que el gen esté accesible para lo que la cromatina debe disminuir su grado de empaquetamiento. La acetilación de determinadas lisinas de las histonas favorece la descondensación de la cromatina. El complejo remodelador de la cromatina o CRM (Chromatin-Remodeling-Machine) tiene afinidad por las lisinas acetiladas de las histonas y se une a ellas a través del dominio llamado "Bromodominio". El CRM desorganiza los nucleosomas aumentando el grado de exposición y de accesibilidad de los promotores. Finalmente el mediador, la ARN polimerasa II y los factores de transcripción hacen que se inicie la transcripción.
El promotor de un gen es una región del ADN con unas características especiales que determina el punto en el que la ARN polimerasa comienza a transcribir un gen. Una vez que la ARN polimerasa ha reconocido el promotor y se ha unido a él en primer lugar se forma el complejo de preiniciación. Este complejo está formado por algunos factores de transcripción y la ARN polimerasa II. Después se forma el complejo de iniciación cerrado al unirse otros factores de transcripción y el mediador. Posteriormente se forma el complejo de iniciación abierto gracias a la actividad helicasa de uno de los factores. En este instante comienza la síntesis de ARNm por la ARN polimerasa II a partir del sitio llamado +1 que marca el punto de inicio de la transcripción de un gen. Hasta que el fragmento de ARNm sintetizado no tiene 8-10 nucleótidos el proceso es reversible siendo posible que el complejo se desorganice y la transcripción del gen no se lleve a cabo. A este fenómeno se le conoce como iniciación abortiva. Cuando ya hay un fragmento de ARN de tamaño adecuado, se fosforila el dominio CTD (Carboxi-Terminal Domain) de la ARN polimerasa II. Esta fosforilación, llevada a cabo por el mismo factor que tiene actividad helicasa o por el mediador, es muy importante, ya que desestabiliza las interacciones que tiene la ARN polimerasa II con algunos factores de transcripción favoreciendo su avance rápido transcribiendo el gen.
• Fase de elongación. Durante la fase de elongación el CTD debe seguir fosforilado. En esta etapa, la ARN polimerasa II cataliza la formación de los enlaces fosfodiéster entre nucléotidos. Intervienen otros factores, conocidos como factores de elongación. Sus funciones son disminuir las pausas de la ARN polimerasa II, desorganizar los nucleosomas y favorecer los procesos de corrección de errores. También intervienen factores de transcripción involucrados en la iniciación. En la elongación se fosforila otra posición del CTD. El CTD fosforilado es reconocido también por proteínas encargadas del procesamiento y maduración del pre-ARNm, de modo que la maduración del pre-ARNm se produce de forma simultánea a la transcripción. Concretamente, la formación de la caperuza (cap) metilada ocurre en la fase de iniciación de la transcripción y normalmente, el proceso de splicing durante la elongación. La poliadenilación comienza en la fase de terminación de la transcripción y acaba una vez finalizada ésta.
• Fase de terminación. El CTD es defosforilado por una fosfatasa. La ARN polimerasa II continúa transcribiendo hasta llegar a una secuencia específica que determina el sitio de poliadenilación. Esta secuencia es reconocida por una endonucleasa. Esta enzima corta el ARNm unos nucleótidos más allá de la secuencia que reconoce, liberándolo. Posteriormente esta secuencia específica es reconocida por una enzima encargada de añadir la cola de poli-A al transcrito.
El proceso de transcripción debe estar finamente regulado, ya que de él depende la adaptación al medio y el buen funcionamiento del organismo. Cada tipo celular y cada estado funcional necesita un perfil de expresión génica diferente que se adapte a sus necesidades. Los factores reguladores pueden incidir sobre cualquiera de los elementos de este complejo proceso. Así hay reguladores que modifican la accesibilidad de los genes, otros que alteran la fase de iniciación, otros que actúan en la fase de elongación y otros en la de terminación. Igualmente existen reguladores de la transcripción que actúan sobre los procesos de maduración del ARN
La regulación de la accesibilidad de los genes depende de las regiones controladoras de locus o LCRs (Locus Control Region), de los “insulators” (aisladores), de los procesos de modificación de histonas, de los procesos de desempaquetamiento y desorganización de nucleosomas y de la metilación del ADN:
• Las LCRs son regiones del ADN donde la expresión génica está favorecida.
• Los “insulators” son regiones del ADN que delimitan a modo de barrera las zonas transcripcionalmente activas de las que están inactivas participando en la correcta organización de la cromatina.
• Las modificaciones en las histonas son muy importantes en el control de la transcripción. La metilación de lisinas impide la transcripción génica ya que las histonas con este tipo de modificación reclutan proteínas silenciadoras de genes que suelen poseer un “cromodominio” que intervienen en la interacción con las histonas metiladas. En cambio, la acetilación de determinadas lisinas favorece la interacción con el CRM que lleva a cabo la desorganización de los nucleosomas y por tanto hace los genes accesibles para ser transcritos.
• La metilación del ADN es otra modificación que produce silenciamiento de genes. La metilación de genes es muy importante en procesos del desarrollo, en la impronta ganética y en el silenciamiento de genes repetidos.
Otro nivel de regulación de la transcripción es el que se lleva a cabo actuando sobre el proceso de iniciación. Existen proteínas activadoras que se unen a elementos próximos al promotor y otras que se unen a los llamados “potenciadores” que pueden localizarse en posiciones más alejadas del promotor. Las proteínas activadoras se unen a sus sitios de unión en el ADN y favorecen el reclutamiento de la ARN polimerasa II y de los factores de transcripción necesarios para que se forme el complejo de iniciación y comience la transcripción. Las proteínas represoras actúan de forma directa uniéndose al operador y evitando que se una la ARN polimerasa o de forma indirecta bloqueando la acción de las proteínas activadoras.
Otros reguladores de la transcripción actúan sobre las proteínas involucradas en el proceso de elongación bloqueando la transcripción a este nivel.
Video complementario
Traducción
Tiene lugar en los ribosomas, de una forma muy similar en procariontes y eucariontes.
Comprende las siguientes etapas:a) Iniciación. Comienza por el triplete iniciador del ARNm (AUG), que está próximo a la caperuza 5'. Este triplete va precedido de la secuencia AGGAGG (secuencia de Shine-Dalgarno ) que es la zona de unión con el ribosoma.
Se forma el complejo de iniciación con los factores de iniciación (FI) y la energía suministrada por el GTP, la subunidad menor del ribosoma reconoce la caperuza y se une al ARNm en la zona proxima al triplete o codón iniciador. Esta caperuza aporta el ARNt iniciador que a su vez aporta el aminoácido metionina. Este ARNt contiene un triplete complementario al AUG, es decir el UAC, llamado anticodón (la proteína sintetizada contiene en su extremo el aminoácido metionina)
Una vez encajado el ARNt-metionina, se liberan los FI y dejan paso a la subunidad mayor del ribosoma, formandose así el ribosoma completo y funcional. En él hay dos sitios claves:
- Sitio P (sitio peptidil) ocupado por el ARNt-metionina
- Sitio A (sitio aminoacil) que está libre para recibir un segundo ARNt (sólo el que su anticodón coincida con el del codón del ARNm) cargado con un nuevo aminoácido.
vídeo complementario
Lipidos
Concepto
Los lípidos son un conjunto de moléculas orgánicas, la mayoría biomoléculas, compuestas principalmente por carbono e hidrógeno y en menor medida oxígeno, aunque también pueden contener fósforo,azufre y nitrógeno, que tienen como característica principal el ser hidrofóbicas o insolubles en agua y sí en disolventes orgánicos como la bencina, el alcohol, el benceno y el cloroformo. En el uso coloquial, a los lípidos se les llama incorrectamente grasas, aunque las grasas son sólo un tipo de lípidos procedentes de animales. Los lípidos cumplen funciones diversas en los organismos vivientes, entre ellas la de reserva energética (triglicéridos), la estructural (fosfolípidos de las bicapas) y la reguladora (esteroides).Características generales
Los lípidos son biomoléculas muy diversas; unos están formados por cadenas alifáticas saturadas o insaturadas, en general lineales, pero algunos tienen anillos (aromáticos). Algunos son flexibles, mientras que otros son rígidos o semiflexibles hasta alcanzar casi una total flexibilidad molecular; algunos comparten carbonos libres y otros forman puentes de hidrógeno.La mayoría de los lípidos tiene algún tipo de carácter polar, además de poseer una gran parte apolar o hidrofóbico ("que le teme al agua" o "rechaza al agua"), lo que significa que no interactúa bien con solventes polares como el agua. Otra parte de su estructura es polar o hidrofílica("que ama el agua" o "que tiene afinidad por el agua") y tenderá a asociarse con solventes polares como el agua; cuando una molécula tiene una región hidrófoba y otra hidrófila se dice que tiene carácteranfipático. La región hidrófoba de los lípidos es la que presenta solo átomos de carbono unidos a átomos de hidrógeno, como la larga "cola"alifática de los ácidos grasos o los anillos de esterano del colesterol; la región hidrófila es la que posee grupos polares o con cargas eléctricas, como el hidroxilo (–OH) del colesterol, el carboxilo (–COO–) de los ácidos grasos, el fosfato (–PO4–) de los fosfolípidos, etc.
Función de los Lipidos
1. Función de reserva. Son la principal reserva energética del organismo. Un gramo de grasa produce 9'4 kilocalorías en las reacciones metabólicas de oxidación, mientras que proteínas y glúcidos sólo producen 4'1 kilocaloría/gr.
2. Función estructural. Forman las bicapas lipídicas de las membranas. Recubren órganos y le dan consistencia, o protegen mecánicamente como el tejido adiposo de piés y manos.
3. Función biocatalizadora. En este papel los lípidos favorecen o facilitan las reacciones químicas que se producen en los seres vivos. Cumplen esta función las vitaminas lipídicas, las hormonas esteroideas y las prostaglandinas.
4. Función transportadora. El tranporte de lípidos desde el intestino hasta su lugar de destino se raliza mediante su emulsión gracias a los ácidos biliares y a los proteolípidos.
Clasificacion de los lípidos
Los lípidos se clasifican en dos grupos, segun su composición en ácidos grasos (lípidos saponificables) o no lo posean (lípidos insaponificables).
- Lípidos saponificables
Acilglicéridos. Cuando son sólidos se les llama grasas y cuando son líquidos a temperatura ambiente se llaman aceites.
Céridos (ceras)
Complejos. Son los lípidos que además de contener en su molécula carbono, hidrógeno y oxígeno, también contienen otros elementoscomo nitrógeno, fósforo, azufre u otra biomolécula como un glúcido. A los lípidos complejos también se les llama lípidos de membrana pues son las principales moléculas que forman las membranas celulares.
Fosfolípidos
Fosfoglicéridos
Fosfoesfingolípidos
Glucolípidos
Cerebrósidos
Gangliósidos
Lípidos insaponificables
Terpenoides
Esteroides
Eicosanoides
- Lipidos simples
Son lípidos simples formados por la esterificación de una,dos o tres moléculas de ácidos grasos con una molécula de glicerina. También reciben el nombre de glicéridos o grasas simples
Según el número de ácidos grasos, se distinguen tres tipos de estos lípidos:
· los monoglicéridos, que contienen una molécula de ácido graso
· los diglicéridos, con dos moléculas de ácidos grasos
· los triglicéridos, con tres moléculas de ácidos grasos.
Los acilglicéridos frente a bases dan lugar a reacciones de saponificación en la que se producen moléculas de jabón.
Ceras
Las ceras son ésteres de ácidos grasos de cadena larga, con alcoholestambién de cadena larga. En general son sólidas y totalmente insolubles en agua. Todas las funciones que realizan están relacionadas con su impermeabilidad al agua y con su consistencia firme. Así las plumas, el pelo , la piel,las hojas, frutos, están cubiertas de una capa cérea protectora.
Una de las ceras más conocidas es la que segregan las abejas para confeccionar su panal.
- Lipidos complejos
Son lípidos saponificables en cuya estructura molecular además de carbono, hidrógeno y oxígeno, hay también nitrógeno,fósforo, azufre o un glúcido. Son las principales moléculas constitutivas de la doble capa lipídica de la membrana, por lo que también se llaman lípidos de membrana. Son también moléculas antipáticas.
En el siguiente vídeo completan la información ya mencionada sobre los lipidos y sus aspectos principales.
A CONTINUACION LOS LIPIDOS EN LA MEMBRANA LIPIDICA

El 98% de los lípidos presente en las membranas celulares son anfipático, es decir que presentan un extremo hidófilo (que tiene afinidad e interacciona con el agua) y un extremo hidrofóbico (que repele el agua). Los más abundantes son los fosfoglicéridos (fosfolípidos) y los esfingolípidos, que se encuentran en todas las células; le siguen los glucolípidos, así como esteroides (sobre todo colesterol).
A CONTINUACION UN VIDEO QUE COMPLMENTA LA INFORMACION
Importancia de los lipidos en la salud
Los lípidos representan alrededor del 30% del total de calorías utilizado día a día por el cuerpo, además de cumplir otras importantes funciones en el organismo como se menciona más adelante. Entre los alimentos que brindan más lípidos al cuerpo tenemos:
- Con lípidos saturados: estos los brindan los alimentos de origen animal, como: mantequilla, carne, grasa, leche, embutidos, entre otros. También se pueden encontrar en aceites como el de coco.
 |
| Ilustración 1 Carne, fuente de lípidos saturados [26] |
- Con lípidos monoinsaturados: provienen de alimentos de origen vegetal como aceite de oliva.
 |
| Ilustración 2 Aceite de Oliva [25] |
- Con lípidos poliinsaturados: dentro de este grupo se encuentra el pescado, aceite de girasol, maní, soya, sésamo.
 |
| Ilustración 3 Aceite de sésamo [27] |
Los lípidos monoinsaturados y poliinsaturados ayudan a disminuir el colesterol dañino ( LDL) y aumentar el beneficioso (HDL), a diferencia de los lípidos saturados que aumentan el colesterol malo.
Medicamentos que presentan lipidos
Se encuentran los hipolipemiantes
Se entiende por hipolipemiante a cualquier sustancia farmacológicamente activa que tenga la propiedad de disminuir los niveles de lípidos en sangre. En elsistema de clasificación anatómica, terapéutica y química, forman un grupo homogéneo denominado C10.
La importancia de estas sustancias viene dada porque el exceso de algunos tipos de lípidos (colesterol o triglicéridos) o de las lipoproteínas es uno de los principales factores de riesgo para la enfermedad cardiovascular, principal causa de muerte en los países desarrollados.
A Continuación se mencionaran algunos
- benfluorex
- benzafibrato
- binifibrato
- clofibrato
- colestipol
- colestiramina
- condroitinsulfúrico ácido
- detaxtrano
- etofibrato
- fenofibrato
- filicol
- gemfibrozilo
- lovastatina
- niceritrol
- nicofibrato
- nicotinato de tocoferol
- pantetina piricarbato
- pirifibrato
- pirozadilo
- pravastatina
- probucol
- simvastatina
- sulodexida
- sultosilato de piperazina
- tiadenol
- tocofibrato
- xilanpolisulfúrico ácido
Presentacion de Algunos hipolipemiantes


FILICOL



miércoles, 1 de junio de 2016
Enzimas:
Conceptos:
Las enzimas son los catalizadores más eficientes que se conocen, Una de
sus principales características es que pueden acelerar una reacción abatiendo
la energía de activación, que es un parámetro de cinética. Los catalizadores no
afectan los aspectos termodinámicos de la reacción. Con pocas excepciones las
enzimas son proteínas o proteínas más cofactores, las enzimas se clasifican en
seis grupos:
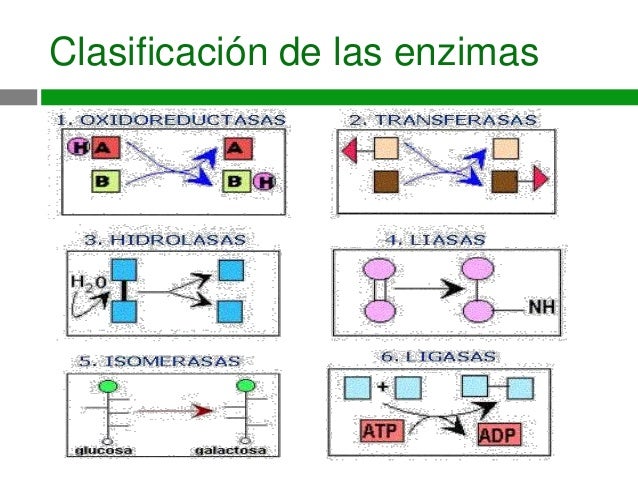
Las enzimas y los sustratos forman complejos enzima-sustrato no
covalentes. En consecuencia, las reacciones enzimáticas son de primer orden en
forma característica respecto a la concentración de la enzima, y en forma
típica muestran una dependencia hiperbólica respecto a la concentración del
sustrato. La hipérbola se describe con la ecuación de Michaelis-Menten.
El primer paso de una reacción catalizada por enzima al sustrato para
formar un complejo. Esto empieza la formación de la especie de estado de transición,
que a su vez forma el producto, el sustrato se une a una porción pequeña de la
enzima llamado sitio activo. Se han propuesto dos modelos para describir la
unión ES: El modelo' de llave y cerradura, en la que la E y el S embonan a la perfección,
y El otro modelo es de Ajuste inducido en el que se considera que a E tiene
flexibilidad conformacional y solo hay un embone perfecto una vez que el S se
ha unido. El sitio activo de la E obliga al sustrato a imitar el estado de transición
de la reacción, que es la forma primaria de abatir la energía de activación de
la reacción.
La Cinética de muchas reacciones catalizadas por enzimas se pueden describir
con el Modelo de Michaelis-Menten, en él ocurre que el complejo ES se dice que
es constante. Podemos aprender mucho sobre la naturaleza de una reacción
catalizada por enzima determinado las constantes cinéticas Km (es la relación
de las constantes de velocidad combinadas por la descomposición de ES divida
entre la constante para su formación) y Kcat (indica la cantidad máxima de
moléculas de sustrato convertidas en producto cada segundo por cada sitio
activo) de la enzima.Cinética de las reacciones con sustratos múltiples:
Estas reacciones pueden seguir un mecanismo secuencial estas consisten
en que todos los sustratos estén presentes para que se libere algún producto
con eventos de unión y liberados de forma ordenada o aleatoria o con un
mecanismo de ping – pong que consiste en liberar un producto antes que se
enlacen todos los sustratos.


Los inhibidores nos brindan mucha información sobre las reacciones
enzimáticas, Un inhibidor es un compuesto que se enlaza con una enzima e
interfiere con su actividad. Existen los inhibidores reversibles son los que se
pueden unir a la enzima y liberarse posteriormente, en cambio un inhibidor
irreversible reacciona con la enzima para producir una proteína que no tiene
actividad enzimática y a partir de la cual es imposible regenerar la enzima
original.
Los tipos de inhibidores
reversibles son:

Los inhibidores irreversibles de enzimas forman enlaces covalentes con
las enzimas y así elimina las moléculas del sitio activo en la población
enzimática. Típicamente, la
inhibición irreversible ocurre por alquilación o acilación de la cadena lateral
de un residuo de aminoácido en el sitio activo.
Los inhibidores reversibles con estructuras que les permitan unirse en
forma específica a un sitio activo son más útiles que los reactivos generales
de sustitución. Estos inhibidores se llaman reactivos dirigidos al sitio
activo, o marcadores de afinidad.
Las Enzimas Alostéricas:

Las enzimas alostéricas son enzimas cuyas propiedades son afectadas por cambios en la estructura. Los cambios estructurales son ocasionados por interacción con moléculas pequeñas. Con frecuencia, las enzimas alostéricas no presentan cinética clásica de Michaelis-Menten debido a su unión cooperativa del sustrato, como el caso de la hemoglobina, que no es enzima.
Muestra una curva de y0 en función de [S] para
una enzima alostérica con enlazamiento cooperativo del sustrato. Las curvas
sigmoides se deben a la transición entre dos estados de la enzima. En ausencia
del sustrato, la enzima está en el estado T. La conformación de cada subunidad
presenta una forma en la que se une ineficientemente al sustrato y la velocidad
de la reacción es baja. A medida que la concentración de sustrato aumenta, las
moléculas de enzima comienzan a unirse al sustrato, aunque la afinidad de la
enzima en el estado T sea baja. Cuando una subunidad se une al sustrato sufre
un cambio de conformación que la convierte al estado R y se efectúa la reacción.
Las propiedades cinéticas de la subunidad enzimática en el estado T y en el
estado R son bastante distintas; cada conformación podría, por sí misma,
exhibir una cinética normal de Michaelis-Menten. El cambio de conformación en
la subunidad que al principio se enlaza a una molécula de sustrato afecta las
demás subunidades en la enzima multisubunidades. Las conformaciones de esas
otras subunidades se desplazan hacia el estado R, donde es mucho mayor su
afinidad hacia el sustrato. Ahora se pueden unir al sustrato a una
concentración mucho menor que cuando se hallaban en el estado T.
Regulación de la actividad enzimática:
La actividad de una enzima regulada cambia como respuesta a señales del
ambiente y permite que la célula responda a las condiciones variables, con
ajuste de las velocidades de sus procesos metabólicos.
Ejemplos de enzimas Alostéricas: La Fosfofructocinasa.
Propiedades generales de las enzimas alostérica:

Dos teorías de la regulación alostérica:
En esta parte utilice estas diapositivas para entender algunas dudas sobre regulación alostérica.
http://es.slideshare.net/cuentadropbox101/regulacin-alostrica
Suscribirse a:
Entradas (Atom)